Amino Acid Transport and Regulation: Mechanisms, Control, and Importance for Cell Function Hours January 5, 2024January 5, 2024
Chirality of Amino Acids: Enantiomers, Differences, and Importance in Biology Hours January 4, 2024January 4, 2024
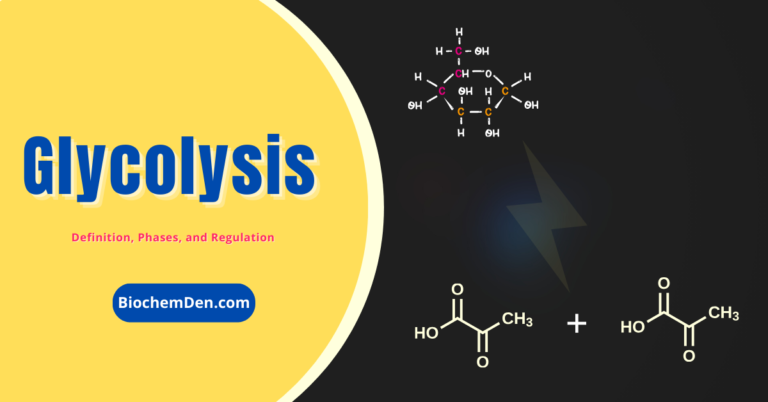
Glycolysis: The Energy-Producing Metabolic Pathway and its Regulation Hours October 29, 2023November 25, 2023
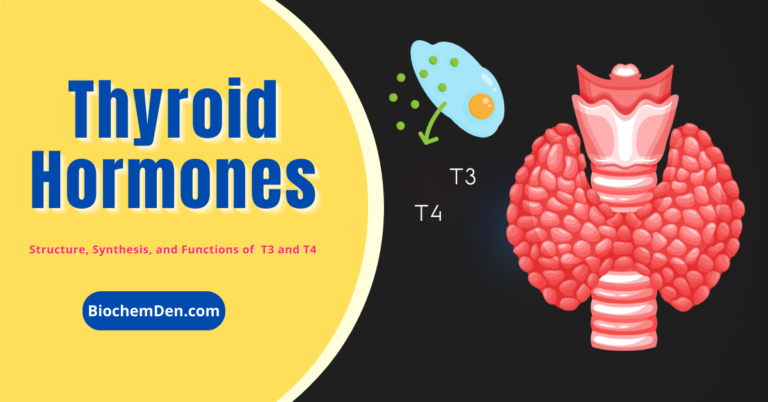
Thyroid Hormones: Structure, Chemistry, Secretions, and Functions Hours October 28, 2023October 26, 2023
Lipids: Structure, Functions and Dietary requirements (Complete Guide) Hours October 26, 2023October 24, 2023